English
English Русский
Русский

Endotoksemiya zamanı sinir sisteminin müxtəlf hissələrində qişa-damar-sinir-qliya elementləri ilə mononukllear faqositlərin qarşılıqlı histotopoqrafiyası və elertron mikroskopik quruluşu
ARA HESABAT
Elmi işlər üzrə prorektor: prof. I.I.İSAYEV
Elmi rəhbər: prof. E.K.QASIMOV
Bakı-2015
İCRAÇILARIN SİYAHISI
Məsul icraçılar:
t.e.d. H.K.Muradov
t.e.n. T.Ə.Sultanova
t.e.n. Y.Ə.Ağayev
İcraçılar:
G.M.Əyyubova.
A.Ə.Əliyarbəyova
N.T.Quliyeva
T.M.Nəcəfova
t.e.n. E.C.Bwnyadzadə
b.e.n. G.Ə.Mehrəliyeva
b.e.n. M.İ.Quliyev
b.e.n. M.R.Mehtiyev
A.İ.Şahməmmədova
A.Ə.Mənsimov
S.H.Nərimanova
A.T.Ibrahimova
İ.O.Əliyeva
G.H.Sadıqova
REFERAT
Mövzu: “Endotoksemiya zamanı sinir sisteminin müxtəlf hissələrində qişa-damar-sinir-qliya elementləri ilə mononukllear faqositlərin qarşılıqlı histotopoqrafiyası və elertron mikroskopik quruluşu”
Tədqiqat obyekti: 20 baş nəzarət qrupu, 10 baş kəskin endotoksemiya modeli yaradımı ağ siçovulların cəsədlərindən götürülmüş baş beyin yarımkürələrinin qabığı, onurğa beyni qanqlionları (OBQ) və oturaq siniri.
İşin məqsədi: OBQ qanqlionunda nero-qlial əlaqələrin və nəzarət qrupu ilə müqayisədə eksperimental ödem zamanı sinir sistemnin müxtəlif hissələrində mononüklear faqositlərin və digər struktur elementlərin histotopoqrafisının və ultrastruktur parametrlərinin dəqiqləşdirilməsidir.
Tədqiqat metodları və aparatlar: 10 ağ siçovulun damar daxili perfuziya üsulu ilə fiksasiya olunduqdan sonra həmin kateterlə 10% jelatində həll olmuş tuş məhlulu aşağı boş venadan xaric olana qədər yeridilərək onurğa beyni qanqlionu (OBQ) onurğa beyni kökləri və sinirləri ilə birgə götürülüb 10% formalin məhlulunda fiksasiya olunmuş Sonra kriostatda dondurularaq 100-150 mkm qalınlığında total kəsiklər alınmışdır. Bundan başqa 10 nəzarət qrupu və 10 baş kəskin endotoksemiya modeli yaradımış ağ siçovulların cəsədlərindən götürülmüş baş beyin yarımkürələrinin qabığı, onurğa beyni qanqlionları (OBQ) və oturaq sinirləri fosfat büferində (pH 7,4) həll edilmiş 1%-li pikrin türşusu, 2,5%-li qlutaraldehid və 2,5%-li paraformaldehid məhlullarında fiksasiya edildikdən sonra ümumi qəbul olunmuş qaydalar üzərə hazırlanmış epon-araldit və spur blokları hazırlanmışdır. Sonunculardan LKB-III, Reichert, və Leica EM UC7 ultratomlarında yarım- və ultranazik kəsiklər alımışdır. Total preparatlar və yarımnazik kəsiklər rəngləngdikdən sonra Latimet (Leika-Almaniya) işıq mikroskopunda baxilaraq lazımi hissələrin şəkilləri Pixera rəqəmsal fotokamerasının (ABŞ) köməkıiyi çəkilmişdir. Ultranazik kəsilər isə 2%-li uranil asetat və 0.4% -li təmiz qurğuşun sitrat məhlulları ilə rəngləndikdən sonra . Hitachi 12E, JEM 1200CX və JEM 1400 tranmission elektron mikroskoplarında tədqiq olunub və lazımı elektronnorqammalar əldə edilmişdir.
Əldə olunan nəticə və yeniliklər: Kəskin endotoksemiya zamanı sinir dəstlərrinin istər epinevral, istərsə də endonevral sahələrdə makrofaqların saylasrının artması illk nəzərə çaropan əlamətlərdin diridir. Endonevral sahədə makrofaqların subperinevral və paravazal sahələrdə klaster şəklində yerləşmələri kəsgin endotoksemiya zamanı bioloji baryerlərdə baş verən dəyişikliklər nəticəsində sinir lifləri ətrafina daxil ola biləcək yad törəmələrin vaxtında kənarlaşdırılması ilə əlaqədar olduğunun göstərircisi kimi qiymətləndirmək olar. Aktivləşmiş makrofaqlar tərəfindən ifraz olunana MİF təsirindən ödem mayesinin miqdarının kəsgin artması nəticəsində mielinli sinir liflərinin müxtəlif dərəcəli degenetativ dəyişikliklərə məruz qalmaları ilə nəticələnir.
Aparılan tədqiqatlar nəticəsində onurğa beyni qanqlionlarının (OBQ) tərkibində yerləşən sinir hüceyrələrinin tun kiçik və açıq böyük olaraq iki qrupa bölünmələrinin heç bir morfoloji əsası olmadığı aşkar edilmişdir. OBQ daxilində neyroqlial əlaqələrin mərkəzi sinir sistemi ilə müqayisədə daha şıx olduqları ultrastruktur səviyyədə dəqiqləşdirilmişdir. Sinir hüceyrələri ilə peyk hüceyrələrinin söykənən hissələrində plazmolemmaları bir-birlərindən ayıran məsafələrin 20-nm az olması onlarların tərkibində yerləşən daşıyıcı, kanal və nasos funkyiyaları daşıyan zulalalar arasında bilavisitə molekulyar əlaqələrin yaranma imkanı imkan yaratmalıdir.
Kəsgin endotoksemiya zamanı beyin qabığının təşkilində iştyirrak edən neyronlarda baş verən dəyişikliklər içərisində nəzərə çarpanı tünd degenerativ neyronları formalaşmasıdır. Maraqlısı odur ki, beyin qabığının hansı qatında yerləşməsinə baxmayaraq tünd neyronların cisimləri və sonunculardan bilavasitə başlanğıc götürən çıxıntıları yaxınlığımda və ya onlarla bilavasitə təmasda olan kəcgin aktivləşmış formaya malik mikroqliya hüceyrəlrinin aşkar edilməsidir. ən səthi qatının formalaşmasında iştirak edən hüdudi neyroqliyanın tərkibində tünd və açıq sotoplamaya malik hüceyrələrin olduğu aşkar edilmışdir. Beləliklə qısa zaman ərzində (cəmi 2 saat müddətində) E.coli endotoksinin təsirindən neyronlarda degenerativ dəyişikliklərin meydana çıxmasında onlara sıxı əlaqələr yaradan mikroqliya hüceyrələrinn mühüm rola malik olduqlqrının göstərir.
Səmərəliyi: Eksperimental ödemlərin və neyrodegeneratriv xəstəliklərin patogenazinin dəqiləşdirilməsi.
Tətbiq etmə dərəcəsi: respublika və beynəlxalq.
Tətbiq etmə sahəsi: Nəzəri və tətbiqi histologiya, nevrologiya və nevropatologiya
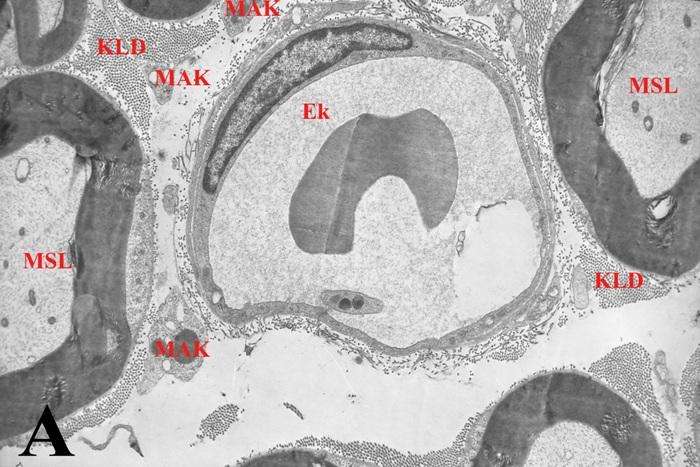
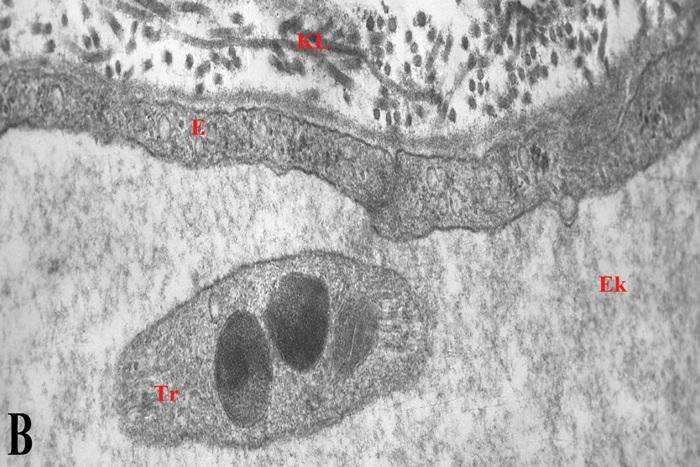
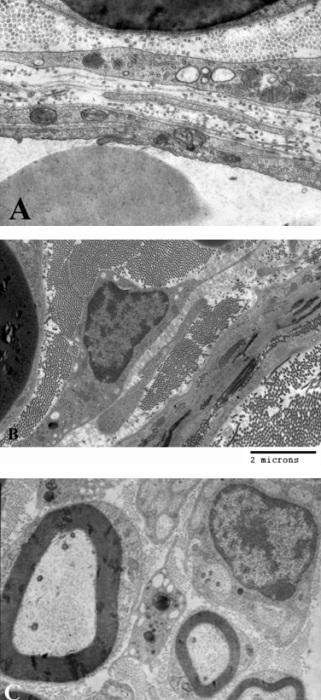
Oldfors (1980), məlumatına görə makrofaqlar endonevrumun birləşdirici toxuma hüceyrələri arasında ən çox sayda olanlarıdır və endonevr sahəsində rast gəlinən hüceyrələrin 2-4 %-ni təşkil edirlər. Kontrol preparatlarda makrofaqlar əsasən endonevr damarların ətrafında (şək. 5.7a), bəzən isə subperinevr sahələrdə (şək. 5.7b) və ya sinir lifləri arasında (şək. 5.7c) yerləşirlər (Qasımov E.K. 2006). Aktivləşməyə məruz qalmamış endonevr makrofaqların nüvə (şək. 5.7b), Holci kompleksi, lizosomlar və müxtəlif ölçülü sitoplazmatik qovuqcuqlar (şək. 5.7c) toplantıları yerləşən hissələrində müəyyən ölçülü qalınlaşmalar müşahidə olunsa da sitoplazmanın əsas hissəsi yastılaşmış çıxıntılar (şək. 5.7a) daxilində yerləşirlər. Kəskin endotoksemiya zamanı makrofaqların topoqrafiq yerləşməsində əsaslı dəyişiklik müşahidə edilmir, ancaq kontrol preparatlarla müqayisədə oturaq sinirin endonevr sahəsində aktivləşmiş makrofaqların miqdarının artması aydın nəzərə çarpır.
Şək. 5.8a-dan göründüyü kimi endonevr sahəsindən venoz qanı epinevr damarlar istiqamətində daşıyan, subperinevr sahədə yerləşən (perinevr qişasından cəmi 4 mkm aralı), perisitar yığıcı venulanın kəsikləri ətrafında bir ədəd makrofaqın nüvə yerləşən mərkəzi hissəsi və ən azı 8 ədəd aktivləşmiş makrofaqların isə periferik hissələrinin fraqmentləri yerləşir. Fraqmentlərin damarların gedişi boyunca həm boylama, həm də köndələn istiqamələrdə yerləşmiş makrofaqlara məxsus olmaları sonuncuların damarətrafında üçölçülü tor əmələ gətirdiklərini göstərir. Qeyd etmək lazımdır ki, makrofaqlar endonevr sahəsində yerləşən digər mikrodamarlarında (məsələn endonevr kapillyarın – şək. 5.8b) ətraflarında bir-biri ilə birləşərək özlərinə məxsus qoruyucu səd yaradırlar. Makrofaqlar bir-biri ilə təmasda olmaqla yanaşı, endonevr sahəsində yerləşən tosqun hüceyrələr və əksərən aktivləşmiş fibrositlərlə də sıx təmasda olur, ya da onlarla muxtəlif şəkilli adhezir əlaqələr (şək. 5.8c) yaradırlar. Əldə etdiyimiz materiallarda rast gəlinən digər makrofaqların ödem mayesinin formalaşmasında iştirak edən damarlarla əlaqəli olduğunu nəzərə alaraq, onlar haqqındakı məlumatlar müvafiq damarlarla birlikdə veriləcəkdir (bax sonraya).
Şək. 5.7. Endonevr sahəsində yerləşən makrofaqın histotopoqrafiyası.
a) kontrol b və c eksperiment. İzahı mətndə verilib.
b) TEM. Böyütmə: a) 27000 b) miqyas 2mkm c) 12600.
Şək. 5.8. Endotoksemiya zamanı endonevr sahəsində damarətrafında
yerləşən makrofaqların histotopoqrafiyası. İzahı mətndə verilib.
TEM. Böyütmə: a) 4320 b) 8640 c) 6480.
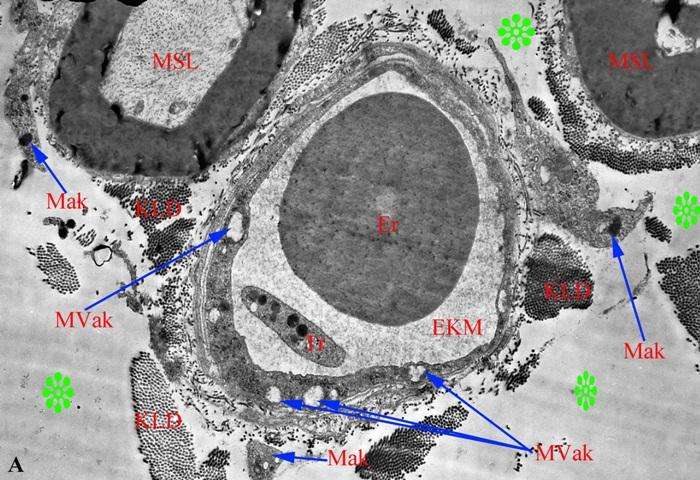
Bununla bərəbər, bizim müşahidələrimizə görə endonevrial sahədə yerləşən kövşək lifli birləşdirici toxumaya aid hüceyrələrdən kapilyarlar ətrafında ən çox rast gələn makrofaqlardır. Şək. 3.19 a-dan göründüyü kimi endonevrial kapilyar ətrafında üç aktivləş miş makrofaqın fraqmenti yerləşmişdir, bəzən hətta bir görmə sahəsində makrofaqın həm boylama, həm də köndələn kəsikləri görünür (şək. 3.20a).
Şək. 3.19. Kəskin endotoksemiya zamanı endonevr kapilyarın mənfəzində və ətrafında yerləşən hüceyrələrin histotopoqrafiyası. İzahı mətndə verilib. TEM. Böyütmə: a) 10560 b) 46200 dəfə.
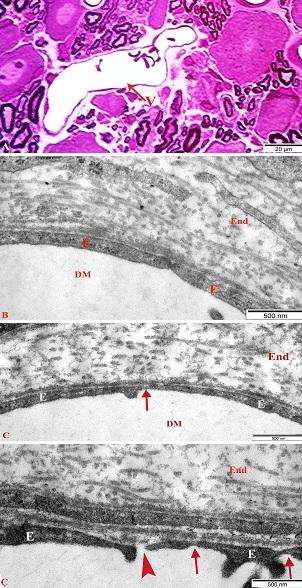
Təsvir edilən endonevrial kapilyarların endotel qatının xeyli hissəsi perisit hüceyrələrinin periferik hissələri ilə örtülməsinə baxmayaraq, birincilərin perisitar örtüyə malik olmayan hissələrində aşkar edilir. Endotel hüceyrələrinin istər lüminal, istərsə də ablüminal səthlərində reaktiv xarakter daşıyan çıxıntılara (protruziyalara) rast gəlinmir. Endotel hüceyrələrinin sitoplazmalarında nəzər diqqəti cəlb edən ikiqat membranla örtülmüş, daraqlarının çox hissəsi destruksiyaya məruz qalmış, vakuola-bənzər mitoxondri qalıqlarının müəyyən edilməsidir. Əgər sonuncuların sayı şək. 3.19-da cəmi iki ədəddirsə, şək. 3.20-də onların sayı altıya çatmışdır.
Bununla yanaşı, şək. 3.20-də təsvir edilən kapilyarın ətraflarında ödem mayesinin yerləşməsinə məxsus olan, tərkiblərində amorf maddəyə belə xarakter olan osmiofillik müşahidə olunmayan sahələrin və tamlıqlarını itirmiş kollagen lifi dəstələrinin sayı şək. 3.19 b ilə müqayisədə xeyli çoxdur.
Digər bir fərq şək. 3.20 a-da nümayiş etdirilən kapilyarın yuxarı sağ tərəfində, damar divarının perisitar örtüyünun olmadığı hissədə, endotel hüceyrəsinin lüminal səthindən cəmi 0,4 mkm məsafədə yerləşən aktivləşmiş markofaqın periferik hissəsinin yerləşməsidir. Bu hissənin 26000 dəfə böyüdülmüş şəklindən aydın görünür ki (şək. 3.20 b), endotellə makrofaqın “təmas” səthində nəinki təsvir edilən hüceyrələrin, hətta yaxınlıqda yerləşən perisitin plazmolemmaları, onları əhatə edən bazal səfhə və sitoplazmatik elementlər nəzərə çarpacaq destruktiv dəyişikliklərə məruz qalmışlar. Sadalanların hamısı endonevrial kapilyarın divarının seçici keçiricilik qabiliyyətinin kəskin pozulduğunu göstərir. Kəskin endotoksemiya zamanı endonevrial kapilyarların mənfəzində onun endotel qatı ilə müxtəlif növ əlaqələr yaradan qanın formalı elementləri aşkar edilir.
Şək. 3.20. Kəskin endotoksemiya zamanı endonevr kapilyarın divarının seçici keçiriciliyinin pozulduğunun elektron-mikroskopik şəkli. İzahı mətndə verilib. TEM. b) a-nın fraqmenti. Böyütmə: a) 9120, b) 19000 dəfə.
OBQ
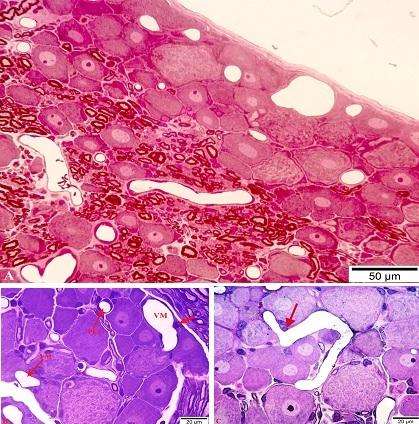
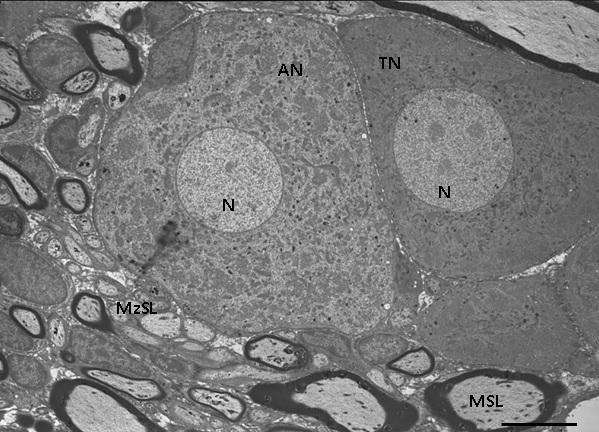
OBQ-də yerləşən sinir hüceyrələrinin sitoplazmasının tündlük dərəcəsinin və bir çox immunhistokimyəvi parametrlərinin tədqiq olunan histoloji kəsiklərin qalınlığından asılılığını [Russo D., 2012] nəzərə alaraq, neyronların rənglənmə dərəcələri ancaq yarım- və ultranazik kəsiklərdə aparılmışdır. Nəticələr göstərir ki, OBQ neyronlarının ancaq 2 qrupa bölünməsi əldə olunmuş faktiki materiallara uyğun gəlmir. Belə ki, şəkil 1A-dan göründüyü kimi, 1 rəqəmi ilə işarə edilmiş neyron görüş sahəsində olan digər neyronlarla müqayisədə ən kiçik ölçüyə (20 μm) malik olsa da, rənglənmə intensivliyinə görə 2 rəqəmi ilə işarə edilən orta diametri 40 μm olan açıq neyrondan, demək olar ki, fərqlənmir. Bununla yanaşı, 3 rəqəmi ilə işarələnmiş 3 ədəd neyronun diametrlərində çox cüzi fərq (37-40 μm) olsa da, onların sitoplazmasının rənglənmə dərəcəsində kəskin fərq aydın görünür. Orta diametrləri təxminən bərabər olan (1-30 μm, 2-31 μm) neyronların birinin tünd, digərinin açıq sitoplazmaya malik olduğu müşahidə edilir (şəkil 1B). Əksər müəlliflərin böyük neyronlar qrupuna aid etdikləri orta diametri 42 μm olan neyronun sitoplazmasının onun ətrafında yerləşən istər kiçik, istərsə də böyük diametrə malik neyronlarla müqayisədə tünd rəngləndiyi nəzər diqqəti cəlb edir (şəkil 1C). 3 rəqəmi ilə göstərilmiş neyronun diametri 2 rəqəmi ilə işarələnən neyronunkundan 1,7 dəfə kiçik olmasına baxmayaraq, rənglənmə intensivliyi demək olar ki, eynidir. 2-ci şəkildə nümayiş etdirilən elektronoqramda köndələn diametrləri bir-birindən çox fərqlənməyən mielinli və mielinsiz sinir lifləri arasında yerləşən neyronlardan birinin tünd, digərinin isə açıq sitoplazmaya malik olduğu heç bir şübhə doğurmur. Həmçinin qeyd etmək lazımdır ki, bu neyronların sitoplazmalarının müqayisəli ultrastrukturu göstərir ki, istər açıq, istərsə də tünd neyronların Nissl elementləri (xromatofil maddə) ilə neyroplazma strukturlarının qarşılıqlı əlaqələrinin kəskin fərqi elektron mikroskopiyanın 30 mindən çox böyüdücüsündə daha qabarıq nəzərə çarpır (şəkil 3A, B).
OBQ neyronlarını ancaq böyük açıq və kiçik tünd qruplara bölən Tandrup T. və həmmüəllifləri 2% halda neyronların bu qrupların heç birinə aid olmadığını göstərirlər [Tandrup T. A, 1993; 1999]. Əldə edilən nəticələr, ələlxüsus eyni diametrə malik, ancaq müxtəlif intensivliklə rənglənən neyronların işıq mikroskopunun 1000 dəfə böyüdülmüş görüş sahələrindən çəkilən mikrofotoların demək olar ki, hamısında aşkar edilməsi yuxarıda şərh edilən məlumatlarla ziddiyyət təşkil edir.
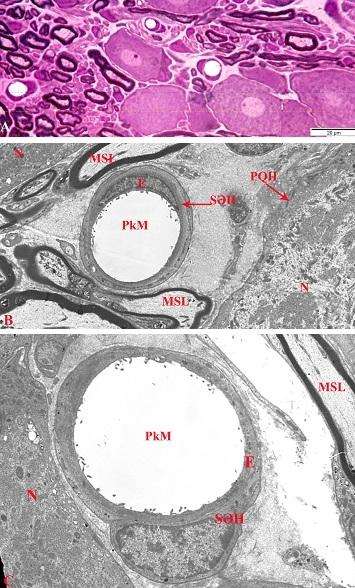
Şək. 1. OBQ-də qliya elementləri və mielinli sinir lifləri ilə əhatə olunmuş müxtəlif intensivlikdə rənglənən psevdounipolyar neyronlar. İzahı mətndə verilmişdir. Ixtisarlar: AN – açıq neyron; TN – tünd neyron; PH – peyk hüceyrələri; MSL – mielinli sinir lifi. Yarımnazik kəsiklər. Boyaq: metilen abısı+Azur II. Miqyas: A-40 μm; B və C-20 μm.
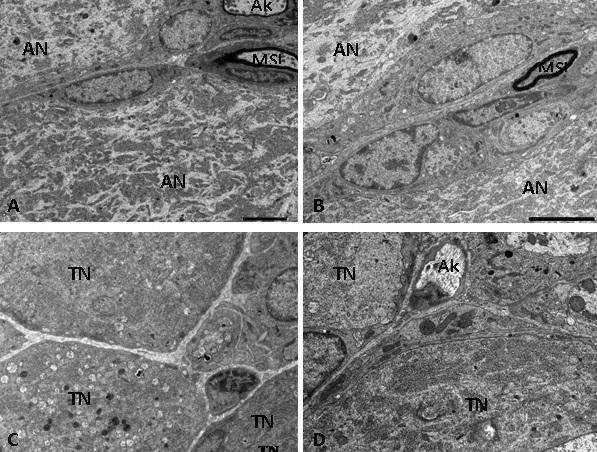
Açıq və tünd neyronun histotopoqrafik vəziyyətlərində hər hansı bir qanunauyğunluq aşkar edilməmişdir. Bununla yanaşı, qeyd etmək lazımdır ki, həm açıq (şək. 4A və B), həm də tünd (şək. 4C və D) neyronların sitoplazmalarında da orqanellərin tərkibində, onların sitoskelet və neyroplazma elementləri ilə qarşılıqlı münasibətlərində nəzərə çarpacaq fərq olduğu aşkar edilmişdir.
Şək. 2. Açıq və tünd psevdounipolyar neyronların elektron-mikroskopik görünüşü. İzahı mətndə verilmişdir. Ixtisarlar: AN – açıq neyron; TN – tünd neyron; N – nüvə; MSL – mielinli sinir lifi; MzSL – mielinsiz sinir lifi. Ultranazik kəsik. Boyaq: uranil-asetat və təmiz qurğuşun-sitrat. Miqyas: 7,4 μm.
Cədvəl 1. Psevdounipolyar neyronların morfometrik parametrlərinin variasion statistik göstəriciləri (n=1030; p=0,95)
|
NEYRONUN PARAMETRLƏRİ |
MİN |
MAX |
X |
S |
SX |
L |
V |
|
Orta diametri (µm) |
11,07 |
69,32 |
30 |
9,8 |
0,31 |
0,64 |
32,67 |
|
Perimetri (µm) |
38,45 |
235,67 |
101,4 |
33,32 |
1,04 |
2,16 |
32,86 |
|
Sahəsi (µm2) |
84,69 |
3358,5 |
683,29 |
507,66 |
15,82 |
32,9 |
74,3 |
|
Forma faktoru (Shape factor) |
0,47 |
0,9 |
0,75 |
0,07 |
0,002 |
0,004 |
8,96 |
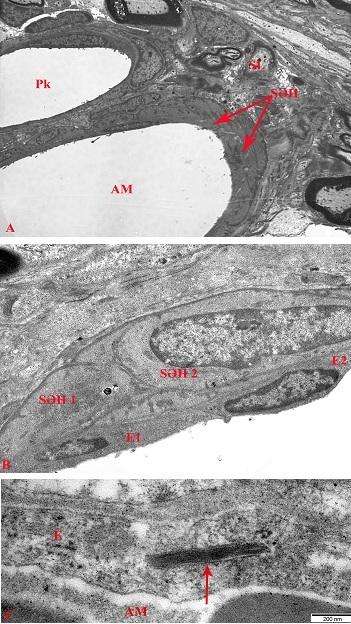
Şək. 3. Açıq (A) və tünd (B) psevdounipolyar neyronların tərkibində orqanellərin, sitoskelet və neyroplazma elementlərinin elektron-mikroskopik şəkilləri. Ixtisarlar: NM – nissl maddəsi; N – nüvə; HA – holci aparatı; M – mitoxondri; SSE – sitoskelet elementləri. Ultranazik kəsik. Boyaq: uranil-asetat və təmiz qurğuşun-sitrat. Miqyas: 500 nm.
Şək. 4. Neyro-qlial əlaqələrə və sitoplazmatik strukturların paylanmasına görə bir-birindən fərqlənən açıq (A, B) və tünd (C, D) psevdounipolyar neyronların elektron-mikroskopik şəkilləri. İzahı mətndə verilmişdir. Ixtisarlar: AN – açıq neyron; TN – tünd neyron; Ak – akson; MSL – mielinli sinir lifi. Miqyas: A, B, C – 5 μm, D – 2 μm.
Yuxarıda göstərilən məlumatları yekunlaşdıraraq qeyd etmək lazımdır ki, istər histoloji, istərsə də elektronmikroskopik səviyyədə OBQ təşkilində iştirak edən neyronların ancaq kiçik tünd və böyük açıq olmaqla 2 qrupa bölünməsinin morfoloji əsası yoxdur.
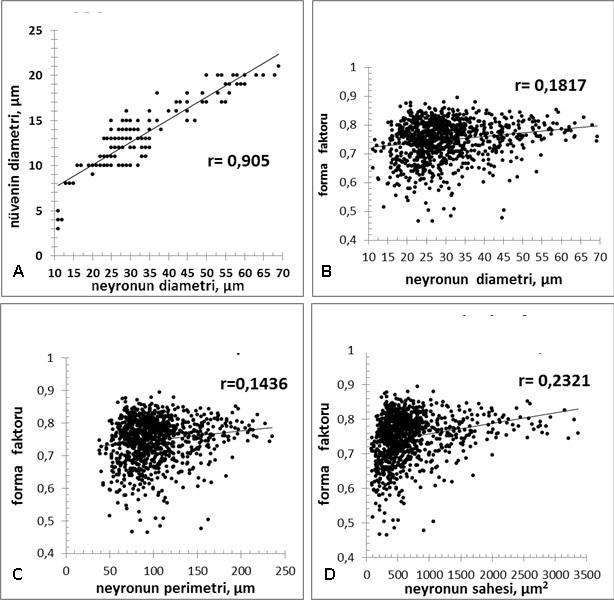
Şək. 5. Psevdounipoiyar neyronların özlərinin və nüvələrinin orta diametrləri (A), birincilərin orta diametri (B), perimetri (C) və sahəsi (D) ilə forma faktoru arasında olan korrelyasion əlaqəni əks etdirən diaqramlar. İzahı mətndə verilmişdir.
Variasiya əmsalının (cədvəl 1) 32-dən yuxarı olması OBQ-nin neyronlarının orta diametrlərinin, perimetrlərinin və sahələrinin bir-birindən kəskin fərqlənən morfoloji parametrlərə malik olduqlarını göstərir. Ancaq gözlənildiyi kimi tədqiq olunan psevdounipolyar hüceyrələrin orta diametri ilə onların perimetri və sahələri arasında güclü korrelyasion əlaqə (r=0,99 və r=0,97) vardır. Bununla birlikdə, neyronların orta diametri ilə həmin neyronların nüvələrinin diametrləri arasında sıx müsbət korrelyasion əlaqə (r=0,905) olduğu aşkar edilmişdir. Göstərilənlərin tam əksinə olaraq, neyronların forma faktorları (shape factor) ilə istər diametri, istər perimetri, istərsə də sahələri arasında korrelyasion əlaqələr 0,14 - 0,23 hüdudunda (şək. 5) tərəddüd edir. Bu isə göstərilən parametrlər arasında korrelyasion əlaqənin demək olar ki, yoxluğunu göstərir. Əgər forma ilə funksiyanın, ümumən qəbul olunmuş, qarşılıqlı əlaqəli olduğunu nəzərə alsaq, forma faktoru ilə psevdounipolyar neyronların orqanel və neyroplazma münasibətləri arasında asılılığın tapılması gələcəkdə psevdounipolyar neyronların yeni morfoloji təsnifatının verilməsinə gətirib çıxara bilər.
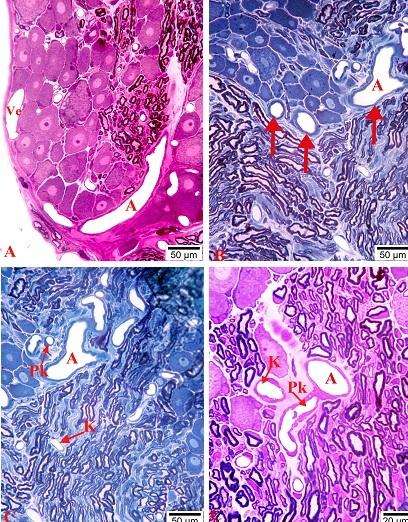
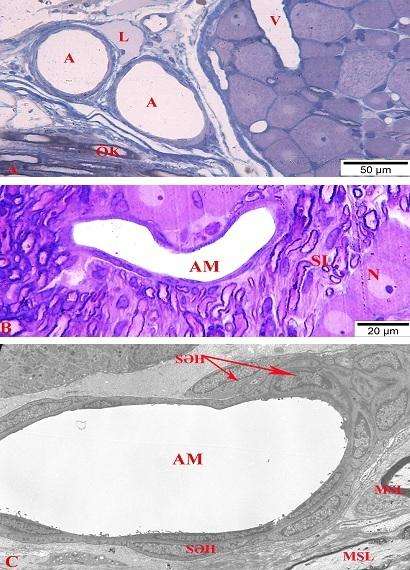
Bəzi müəlliflər ağ siçovullarda kapsulu dəlib qanqliona daxil olan damarı arteriya kimi təsvir edirlər. Ancaq istər kapsulun bilavasitə yaxınlığında (şək. 4.23 a), istərsə də qanqlion daxili arterial damarların divarlarında cəmi bir (şək. 4.23 b, c), bəzən isə yarımçıq iki qat saya əzələ hüceyrələrinin yerləşməsi (şək. 4.24 a və b) və endotel hüceyrələrinin sitoplazmalarında multitubulyar (Weibel-Palade) cisimciklərinin aşkar olunması (şək. 4.24 c-də oxla göstərilib) onların arteriola olduqlarını bir mənalı təsdiq edir.
Şək. 4.23. Onurğa beyni qanqlionu yaxınlığında yerləşən arterial, venoz və limfa damarlarının mikroskopik şəkilləri (A). Kapsul daxili arteriolanın və onun ətrafında yerləşən strukturların histoloji (B), elektron mikroskopik (C) şəkilləri. A, B yarımnazik kəsiklər, C elektronoqramma. İzahı mətndə verilmişdir. Boyaq: A metilen abısı, B metilen abısı / azur II + qələvi fuksin, C uranil asetat və təmiz qurğuşun sitrat. Böyütmə: miqyas A) 50 mkm, B) 20 mkm, C) 3000 dəfə.
Şək. 4.24. Onurğa beyni qanqlionu endonevral arterial damarların və onların ətrafında yerləşən strukturların elektron mikroskopik şəkilləri. TEM. İzahı mətndə verilmişdir. Boyaq: uranil asetat və təmiz qurğuşun sitrat. Böyütmə: A) 2400 dəfə, B) 12000 dəfə, C) miqyas 200 nm.
OBQ daxilində kapsulun yaxınlığında yerləşən arteriolaların divarlarının bir qədər qalın olmasının səbəbi kapsulun daxili qatının təşkilində iştirak edən ixtisaslaşmış hüceyrələrin onları müəyyən məsafədə müşahidə etmələri ilə əlaqədardır. Şək. 4.23 a-da verilən məlumatlara (bax yuxarı) əlavə olaraq onu qeyd etmək lazımdır ki, kapsuldan bir qədər aralı köndələn kəsiyi görünən arteriola (yuxarı solda) kapsul yaxınlığında çəp istiqamət alaraq OBQ-nin kapsulunun daxili qatına yaxınlaşır (kapsul və ön kök qişası arasında). Təsvir olunan yarımnazik kəsikdə digər diqqəti cəlb edən bir tərəfdən arteriolanın kəsikləri arasında xarakterik topoqrafik vəziyyətdə yerləşən limfa kapilyarının mövcudluğu (L ilə işarə olunub), digər tərəfdən isə sinir hüceyrələrinin cisimləri arasında arteriolalarla müqayisədə nazik divara malik, yastılaşmış venulaların kəsiklərinin görünməsidir (şək. 4.23 a).
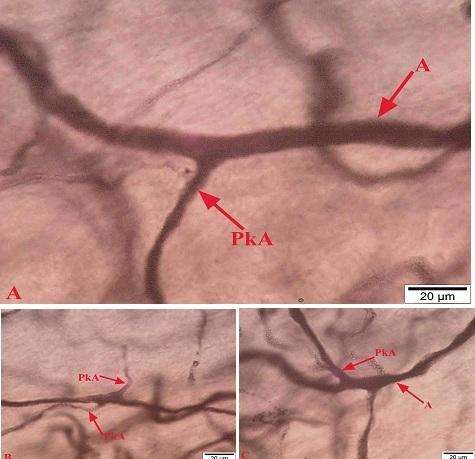
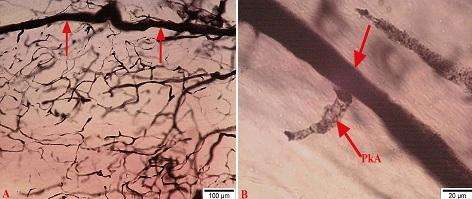
Kapsulu dələrək OBQ-yə daxil olan arteriolalar ilk olaraq qalxan və enən istiqamətdə diametrcə kiçik olmayan şaxələr verirlər (şək. 4.25 a-da oxlarla göstərilib). Göstərilən şaxələr digər mənbələrdən ayrılan arteriollarla uc-uca birləşməklə yanaşı gedişləri boyu yan şaxələr verərək müxtəlif sıxlığa malik orqandaxili subkapsulyar damar torunu əmələ gətirirlər.
Şək. 4.25. Perfuziya üsulu ilə fiksasiya edildikdən sonra qalxan aorta daxilinə yeridilmiş tuş məhlulu (10% li jelatində həll olmuş) ilə aşkarlanmış onurğa beyni qanqlionunun kapsul daxili arterial damarlarının mikroskopik şəkilləri. Kriostat kəsikləri. İzahı mətndə verilmişdir. Miqyas A 100 mkm, B 20 mkm.
Subkapsulyar şaxələri verdikdən sonra arteriolaların davamı sinir hüceyrələrinin qlial örtükləri və ya sinir liflərinin arasında əsasən boylama (şək. 4.25 b) və qövsü istiqamətlərdə (şək. 4.26 a və b) gedərək müxtəlif səthlərə doğru şaxələr verirlər. Qanqlion daxili arteriolaların qövsü gedişə malik olmalarının göstəricilərindən biri, eyni bir yarımnazik kəsikdə onların müxtəlif formalı profillərinin görünməsidir (şək. 4.26 b-də oxlarla nişanlanıblar). Kapsul daxili damar toruna daxil olan arteriolalar öz gedişləri boyu və sonda divarlarının təşkilində yığılma qabiliyyətinə malik hüceyrələr olan prekapliyar arteriolalara başlanğıc verirlər (şək. 4.26 c və ç).
Qanqliondaxili arteriolalardan həm kiçik diametrli arteriolalar (şək. 4.27 a), həm də prekapilyar arteriolalar öz mənbələrindən T hərfini xatırladan düz bucaq altında ayrılırlar (şək. 4.28 a, b, c). Təsvir olunan yerlərin digər xarakter xüsusiyyəti kiçik diametrli arteriolaların ayrılan yerlərində differensiasiya etmiş saya əzələ hüceyrələrinin, prekapilyarların ayrıldıqları yerlərin ətrafında isə primitiv saya əzələ hüceyrələrinin yerləşmələri üçün dairəvi çöküklüklərin aşkar edilməsidir.
Sonuncu xüsusiyyət magistral arteriolalardan ayrılan kiçik diametrli arteriolaların demək olar ki, bütün gedişləri boyu daha aydın nəzərə çarpır (şək. 4.27 b, c, d, e, f). Əksərən prekapilyar arteriolaların başlanğıc hissəsində onların mənfəzinin diametri 4-5 mkm-dən artıq olmur (şək. 4.29 a).
Prekapilyar arteriolaların əsas xarakter xüsusiyyətləri aşağıdakılardır:
- diametri kiçik olmasına baxmayaraq onların mənfəzlərinin 3 və ya 4 endotel hüceyrələri ilə əhatə olunması (müqayisə üçün demək olar ki, eyni diametrli kapilyar damarlarda endotel hüceyrələrinin sayı 1-3-dən artıq olmur);
- endotel hüceyrələrinin istər nüvə yerləşən mərkəzi hissələri, istərsə də periferik hissələri kapilyarlarla müqayisədə 1,5-2 dəfə qalın olur (şək. 4.30 a və b);
- arteriollardan fərqli olaraq prekapilyar arteriolaların subendotel sahələrində elastiki səfhə qalıqlarına təsadüf edilmir (şək. 4.29 b və c);
- prekapilyar arteriolaların periendotelial hüceyrələri öz ultrastruktur quruluşlarına görə perisitlərlə differensiasiya etmiş saya əzələ hüceyrələri arasında keçid olduğunu nəzərə alaraq onları primitiv saya əzələ hüceyrələri adlandırmaq daha məqsədə uyğundur (şək. 4.29 b, c və sək. 4.30 a); - prekapilyar arteriolalar arteriollardan ayrıldıqdan sonra mənfəzlərinin diametri iki dəfə artsa da ultrastruktur olaraq onların divarının təşkilində iştirak edən hüceyrəvi elementlərinin quruluşlarında nəzərə çarpacaq dəyişikliklər müşahidə edilmir (şək. 4.29 c);
Şək. 4.26. Onurğa beyni qanqlionu kapsul daxili damar torlarının formalaşmasında iştirak edən arteriolaların gedişi və şaxələnmə tiplərinin mikroskopik şəkilləri. Yarımnazik kəsik. İzahı mətndə verilmişdir. Boyaq: A, Ç metilen abısı / azur II + qələvi fuksin, B, C metilen abısı. Miqyas A, B, C 50 mkm, Ç 20 mkm.
Şək. 4.28. Perfuziya üsulu ilə fiksasiya edildikdən sonra qalxan aorta daxilinə yeridilmiş tuş məhlulu (10% li jelatində həll olmuş) ilə aşkarlanmış onurğa beyni qanqlionunun kapsuldaxili damar torunda prekapilyar arteriolaların formalaşma tiplərinin mikroskopik şəkilləri. Kriostat kəsikləri. İzahı mətndə verilmişdir. Miqyas hər üçündə 20 mkm.
Şək. 4.29. Onurğa beyni qanqlionu kapsul daxili prekapilyar arteriya və onun ətrafında yerləşən strukturların mikroskopik (A), ultrastruktur (B, C) şəkilləri. A yarımnazik kəsik, B, C elektronoqrammalar. İzahı mətndə verilmişdir. Boyaq: A metilen abısı / azur II + qələvi fuksin, B, C uranil asetat və təmiz qurğuşun sitrat. Böyütmə: A) miqyas 20 mkm, B) 2700 dəfə, C) 6000 dəfə.
- prekapilyar arteriolaların kapilyara keçdiyi nahiyələrindən boylama və çəp istiqamətdə keçən kəsiklərin öyrənilməsi göstərir ki, keçid zonalarında endotel hüceyrələri ətrafında perisit hüceyrələri ilə birlikdə primitiv saya əzələ hüceyrələrinin yerləşdiyi aydın görünür (şək. 4.30 a və b);
Şək. 4.30. Onurğa beyni qanqlionunda prekapilyar arteriolanın başlanğıc hissəsinin (A) və endotel hüceyrələri arasında əlaqələrin (B) elektron mikroskopik şəkilləri. TEM. İzahı mətndə verilmişdir. Boyaq: uranil asetat və təmiz qurğuşun sitrat. Böyütmə: A) 10000 dəfə, B) 42000 dəfə.
Beləliklə, primitiv saya əzələ hüceyrələri prekapilyar arteriolaların bütün gedişi boyu aşkar edilirlər;
- primitiv saya əzələ hüceyrələrinin ultrastruktur olaraq nəzərə çarpan xüsusiyyətləri istər nüvə ətrafı, istərsə də periferik hissələrdə perisitlərlə müqayisədə mitoxondrilərin sayının xeyli artmasıdır (şək. 4.30 a);
- maraqlısı odur ki, mitoxondrilər primitiv saya əzələ hüceyrələrində mərkəzi hissələri tutduqları halda həmin hüceyrələrin plazmolemmalarına yaxın hissələrində isə aktin-miozin kompleksinin və sitoskelet elementlərinin üstünlük təşkil etməsidir. Ona görə də differensiasiya etmiş saya əzələ hüceyrələri ilə müqayisədə nisbətən açıq (şək. 4.30 a), ancaq perisitlərlə müqayisədə isə tünd zolaq şəkilli sahələr aydın görünür.
Son olaraq onu qeyd etmək lazımdır ki, OBQ-də yerləşmiş sinir hüceyrələrinin və onu əhatə edən qlial elementlərin qidalanmasını arteriolalar arasında müşahidə olunan anastomozlar və yerli qan təchizatı miqdarının tənzimləyicisi strukturuna malik olan kapilyarönü damarlar - prekapilyar arteriolalar təmin edir.
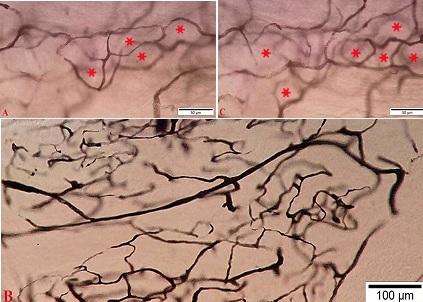
Prekapilyar arteriolalardan başlayan diametri 4-10 mkm arasında tərəddüd edən damarların əksəriyyəti kapilyarlara aiddirlər. OBQ daxilində kapilyarların iştirakı ilə formalaşan damar torlarının xarakter xüsusiyyətləri əsasən onların qanla təchiz etdikləri strukturların tərkibindən asılıdır. Belə ki, mielinli və mielinsiz sinir liflərinin üstünlük təşkil etdikləri hissələrdə qan kapilyarlarının gedişi əsasən boylama xarakter daşıdığı halda (şək. 4.31 b-nin mərkəzi hissəsində), sinir-qliya cütlüyü yerləşdiyi hissələrdə onlar ilgəkşəkilli gedişə malik olub (şək. 4.31 a və c) ilk baxışdan bir və ya bir neçə sinir-qliya cütlüyünü hər tərəfdən əhatə edən (şək. 4.31 a və c-də ulduzlarla göstərilib) halqa şəkilli tor formalaşdırırlar.
Ancaq diqqətlə baxdıqda aydın görünür ki, halqanın formalaşmasında iştirak edən kapilyarların bir hissəsi dumanlı görünüşə malik olurlar. Bu isə bir tərəfdən halqanın qapanmadığını, ikinci tərəfdən isə ümumiyyətlə görünənlərin eyni səthdə yerləşmədiklərini və hər bir qliya-sinir cütlüyünün ətrafında səbətə bənzər damar torunun formalaşmadığını göstərir. Ultrastruktur olaraq OBQ kapsul daxili damar torunun tərkibində aşkar edilən kapilyarların divarlarının təşkilində endotel hüceyrələri ilə yanaşı, bu və ya digər dərəcədə perisitlər iştirak edirlər (bax sonraya).
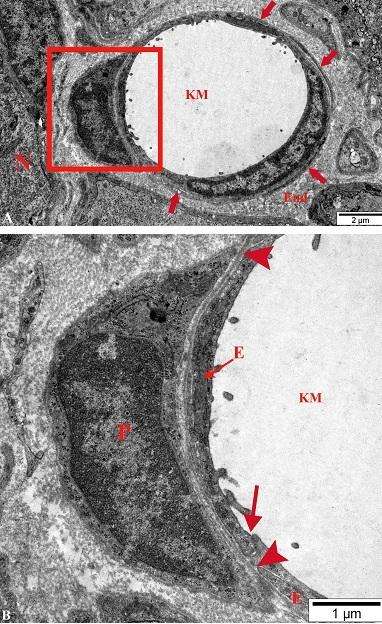
Şək. 4.32 a-da dörd ədəd sinir hüceyrələrinin qlial örtükləri, mielinsiz sinir lifləri, kollagen lifləri dəstələri və fibrositlərin periferik hissələrini iştirakı ilə formalaşan endonevral örtüklə əhatə olunmuş kapilyar nümayiş etdirilir. Üç ədəd endotel hüceyrələrinin ancaq birinin nüvə yerləşən mərkəzı hissəsi, onun və digər iki endoteliositlərin periferik hissələri bir-birləri ilə sıx əlaqələrlə (biri şək. 4.32 b-də oxla göstərilib) birləşərək damar mənfəzini hər tərəfdən əhatə edirlər. Onların lüminal səthləri ilə birləşmiş və sərbəst görünən barmaqvari çıxıntılarının müxtəlif formalı profilləri görünür. Elektronoqrammanın sol tərəfində peristin mərkəzi hissəsinin yerləşdiyi yerin böyüdülmüş fraqmenti şək. 4.32 b-də verilmişdir.
Şək. 4.31. Perfuziya üsulu ilə fiksasiya edildikdən sonra qalxan aorta daxilinə yeridilmiş tuş məhlulu (10% li jelatində həll olmuş) ilə aşkarlanmış onurğa beyni qanqlionu kapsuldaxili sahədə yalançı unipolyar sinir hüceyrələri ətrafı formalaşan damar toru (A, C) ilə sinir lifləri yerləşən hissənin qidalanmasında iştirak edən damar toru (B) ilə müqayisəli xarakteristikası. Kriostat kəsikləri. İzahı mətndə verilmişdir. Miqyas A, C 50 mkm, B 100 mkm.
Şək. 4.32. Onurğa beyni qanqlionunda periendotel sahəsində perisitlərin mərkəzi hissələri yerləşən endonevral kapilyar və onun ətrafında olan strukturların elektron mikroskopik şəkilləri. B A-nın fraqmentidir. TEM. İzahı mətndə verilmişdir. Boyaq: uranil asetat və təmiz qurğuşun sitrat. Miqyas A 2 mkm, B 1 mkm.
Göründüyü kimi, nüvə dəlikləri səviyyələrindən başqa, nüvə ortüyü ilə sıx əlaqədə olan və mərkəzdə yerləşən euxromatin strukturları arasında tünd toplantılar şəklində görünən heteroxromatinin üstünlük təşkil etdiyi nüvə perisitin mərkəzi hissəsini demək olar ki, təklikdə doldurur. Onun ətrafinda nazik zolaq şəklidə görünən sitoplazmanın tərkibində sərbəst ribosomlardan və sitoskelet elementlərindən başqa digər strukturlar aşkar edilmir. Perisiti hər tərəfdən əhatə edən bazal səfhənin endoteliositlərin bazal səfhələri ilə birləşərək üç qatlı bazal zar əmələ gətirmələri şək. 4.32 b-də ox başları ilə göstərilmişdir. İlk baxışdan təsvir olunan kapilyarın ancaq şək. 4.32 b-də nümayiş etdirilən hissəsinin perisitar örtüyə malik olduğu fikri yaranır, ancaq şək. 4.32 a-ya diqqətlə baxdıqda, daha etibarlısı kompyuterdə miqyası böyütdükdə endotel hüceyrələrinin çox hissəsinin perisitlərin periferik hissələri ilə örtüldükləri (oxlarla nişanlanıblar) aşkar edilir.
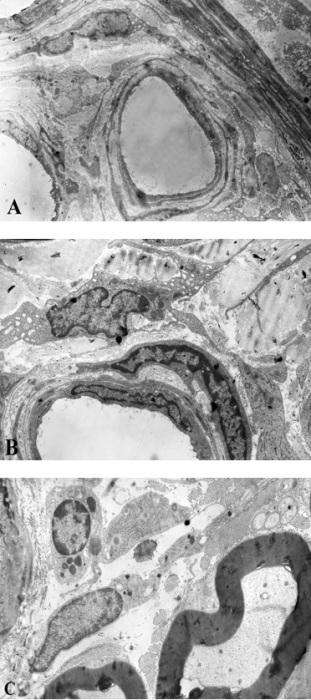
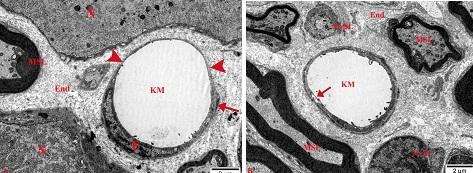
Yuxarıda verilənlərin tam əksi olaraq, şək. 4.33 a-da nümayiş etdirilən kapilyarın ancaq cüzi bir hissəsinin perisitin periferik hissəsi ilə əhatə olunduğunu görürük. Elektronoqrammada (şək. 4.33 a) digər nəzər diqqəti cəlb edən fakt endoteliositlərin periferik hissələrinin hədsiz nazikləşdiklərindən köndələn ölçülərinin (sərhədləri ox başları ilə nişanlanıb) 125 nm-dən artıq olmamalarıdır.
Şək. 4.33. Onurğa beyni qanqlionunda endotel qatı hissəvi olaraq perisit çıxıntıları ilə əhatə olunmuş kapilyarlar və onların əhatəsində olan strukturların sinir hüceyrələri (A) və mielinli, mielinsiz sinir lifləri (B) ilə qarşılıqlı histotopoqrafiyası. Elektronoqrammalar. TEM. İzahı mətndə verilmişdir. Boyaq: uranil asetat və təmiz qurğuşun sitrat. Miqyas hər ikisində 2 mkm.
Şək. 4.33 b-də nümayiş etdirilən kapilyar bir neçə fərqli xüsusiyyətə malikdir: birinci - ətrafinda endonevral strukturlarla yanaşı ancaq mielinli və mielinsiz sinir lifləri yerləşirlər; ikinci – divarının təşkilində həm endoteliositlərin, həm də perisitlərin ancaq periferik hissələri iştirak edirlər, üçüncü – endotel hüceyrəsinin periferik hissəsində makropinositoz əlaməti (oxla göstərilib) müşahidə olunur.
Beləliklə, OBQ kapsul daxili damar torunun tərkibində endoteliositlər və perisitlərin strukturu və qarşılıqlı əlaqələrində fərqli xüsusiyyətlərə malik kapilyarların olduğunu görürük. Endonevral sahədə kapilyarlar demək olar ki, sərhədsiz olaraq postkapilyar venulalara keçirlər. Əvvəlcədən qeyd etməliyik ki, OBQ daxilində yerləşən venoz damarların diametrlərinin bəzən 55 mkm olmasına baxmayaraq (şək. 4.12) divarlarının ancaq bir qat endotel və yarımçıq perisit hüceyrələr qatından təşkil olunduğunu nəzərə alaraq onları vena yox, venulalar adlandıracağıq [215].
Əldə olunan məlumatlar göstərir ki, istər kapilyarların, istərsə də onların birləşməsi nəticəsində formalaşan postkapilyar venulaların gedişləri boyu onların mənfəzlərinin genişlənmiş və daralmış hissələri bir-birini əvəz edir (şək. 4.34).
Şək. 4.34. Onurğa beyni qanqlionunda kapsul daxili damar torunda yerləşən kapilyar və postkapilyar venulaların gedişi boyu formalaşan daralmaların mikroskopik şəkli. Şəkil kompyuterin fotoşopun inversiya proqramı vasitəsilə işlənilmişdir. Kriostat kəsiyi. İzahı mətndə verilmişdir. Miqyas 50 µm.
Aparılan morfometrik ölçülərin nəticələrinə görə, şək. 4.34-də sağ tərəfdən başlayaraq sola doğru ardıcıl olaraq oxla göstərilmış kapilyarın diametri 4, 12, 4, 6, 5, 12 mkm və onun ətrafda yerləşən kapilyarlarla birləşməsindən formalaşan postkapilyar venulanın başlanğıc hissəsinin diametri 13 mkm olduğu halda, axırıncı oxla nişanlanmış hissəsində onun diametri cəmi 6,5 mkm-ə bərabərdir.
Venulaların gedişi boyu yerləşən daralmalar nahiyəsində perisit, bəzən isə primitiv saya əzələ hüceyrələrinin köndələn kəsikləri aşkar edilir (şək. 4.35 b-də oxla göstərilib). Qeyd etmək lazımdır ki, bəzən təsvir olunan daralmalara kapilyarların postkapilyar venulaya keçdikləri nahiyələrdə də rast gəlinir.
Şək. 4.35. Onurğa beyni qanqlionu kapsul daxili venoz damarlar və onları əhatə edən strukturların mikroskopik şəkilləri. Yarımnazik kəsiklər. İzahı mətndə verilmişdir. Boyaq: metilen abısı / azur II + qələvi fuksin. Miqyas A 50 mkm, B, C 20 mkm.
Şəkil 4.35 c-də nümayiş etdirilən yarımnazik kəsikdə 9 mkm diametrinə malik kapilyarın diametri 14 mkm olan postkapilyar venulaya keçdiyi yerdə perisit hüceyrəsinin köndələn kəsiyi (oxla göstərilib) aydın görünür.
Venulaların gedişi boyu rast gəlinən daralmaların ətrafında perisitlərin köndələn istiqamətdə yerləşmələrini ultrastruktur olaraq şək. 4.36 a və b-də nümayiş etdirilib. Göründüyü kimi, elektronoqrammaların hər ikisində oxla nişanlanmış perisitlərin mərkəzi hissələri yerləşən nüvələr və onların ətrafında sitoplazmatik strukturlar ancaq nazik zolaq şəklində görünürlər.
Əgər perisitlər boylama və çəp istiqamətlərdə yerləşsəydilər onların periferik hissələrinin profilləri təqdim olunan elektronoqrammalarda aşkar edilməli idilər.
Kapilyar və venulaların genişlənmiş hissələrinin mikrodamarların mübadilə səthlərinin artırmasına xidmət etdikləri halda, təsadüf edilən daralmaların ətrafında yığılma qabiliyyətinə malik hüceyrələrin yerləşməsi onların qanqlion daxili qan cərəyanının tənzimində iştirak etdiklərini söyləməyə əsas verir.
Perfuziya üsulu ilə fiksasiya olunmuş qanqlionların yarımnazik kəsiklərində aparılan morfometrik ölcülərin və damarların divarlarında iştirak edən hüceyrələrin tərkibi göstərir ki, OBQ daxilində yerləşən damarların böyük əksəriyyəti venulyar damarlara aiddirlər.
Maraqlısı odur ki, şək. 4.35 a-da nümayiş etdirilən yarımnazik kəsikdə olan 18 mikrodamardan 12-si, sək. 4.35 b-də 7-dən 4-ü, şək. 4.35 c-də 5-dən 4-ü mənfəzlərinin diametrinə görə postkapilyar və yığıcı venulalardır.
Kapilyar torundan müxtəlif üsullarla başlayan (şək. 4.34), diametrləri 11 mkm-dən 15 mkm-ə qədər tərəddüd edən postkapilyar venulalar OBQ-nin subkapsulyar səhəsində bir-birləri ilə birləşərək diametrləri 30-55 mkm-ə bərabər olan yığıcı venulaları formalaşdırırlar. Sonuncular OBQ-nin kapsulunu çəpinə dələrək orqan xarıci venoz damarlara açılırlar (şək. 4. 11).
Venaların subkapsulyar sahədə yerləşmələri və qanqlion kapsulunu çəpinə dəlməsi qanqliondaxili təzyiqin təsirindən postkapilyarda müqaviməti artırıb-azaltmaqla OBQ daxili hidrostatik təzyiqin lazımi səviyyədə saxlanılmasını təmin edir.
Şək. 4.36. Onurğa beyni qanqlionu kapsul daxili damar torunda rast gəlinən daralmaların və onların ətrafında yerləşən perisitlərin köndələn kəsiklərinin elektron mikroskopik şəkilləri. TEM. İzahı mətndə verilmişdir. Boyaq: uranil asetat və təmiz qurğuşun sitrat. Böyütmə: A) 4500 dəfə, B) 4200 dəfə.
OBQ kapsul daxili damar toru üçün xarakter olan əlamətlər içərisındə istər struktur, istərsə də funksional cəhətcə diqqət yetirilməli olan kapilyarların və əsasən növündən asılı olmayaraq bütün venulaların daxili qatlarını örtən endoteliositlərin periferik hissələrinin qalınlıqlarının hədsiz nazik olmalarıdır. Sonuncu əlamət, kapilyar və venulaların divarlarının ancaq endotel hüceyrələrinin periferik hissələrindən təşkil olunmuş fraqmentlərinin işıq mikroskopunda da çəkilmiş şəkillərində (şək. 4.35 c və 4.37 a) tam aydınlığı ilə nəzərə çarpırlar.
Elektron mikroskopik olaraq endoteliositlərin periferik hissələri ya mütəmadil olaraq nazikləşərək sonda 47 nm-ə bərəbər olur (şək. 4.37 b-də oxla göstərilib), ya da periferik hissələrin gedişi boyu qalınlığı 42 nm-dən artıq olmayan buğum şəkilli daralmalar meydana çıxır (şək. 4.37 c-də oxla göstərilib).
Maraqlı olan OBQ kapsul daxili kapilyar və venulaların endotelial hüceyrələrinin periferik hissələrinin nazikləşmiş fraqmentlərində tək bir kaveolanın iştirakı ilə transendotelial kanalın formalaşma imkanının aşkar edilməsidir (şək. 4.38 a-da oxla göstərilib). İkinci bir tərəfdən çox nüdurət olsa da (elektron mikroskopik olaraq tədqiq edilmiş 170 damar profilindən cəmi birində) venulaların divarının təşkilində iştirak edən endoteliositlərin periferik hissəsində iki ədəd diafraqmalı (şək. 4.37 ç-də oxlarla göstərilib) və bir ədəd diafraqmasız pəncərənin (fenestranın) aşkar edilməsidir (ox başı ilə göstərilib). Göstərildiyi kimi, damar torunun təşkilində iştirak edən damarlar içərisində nüdurət halda pəncərəli endotelə rast gəlinsə də, OBQ kapsul daxili torunun təşkilində iştirak edən damarların növündən asılı olmayaraq hamısı daxildən somatik tipli endoteliositlərlə örtülürlər.
Şək. 4.38 b-də kapsul daxili damarlardan arteriolanın endotel hüceyrələri arasında təsadüf edilən əlaqə formaları nümayiş etdirilir. Oxlarla hüceyrəarası əlaqənin başlanğıc hissəsində (solda) və təxminən ortasında qonşu endotel hüceyrələrinin plazmolemmalarının zarlı qatlarının bitişmələri nəticəsində formalaşan sıx əlaqə, ox başı ilə isə mexaniki əlaqə formalarına aid edilən desmosom göstərilmişdir.
Şək. 4.38 c-də kapsul daxili prekapliyar arteriolanın endotel hüceyrələri arasında 5-dən artıq bitişmə nöqtələri olan sıx əlaqə nümayiş etdirilmişdir.
Şək. 4.37. Onurğa beyni qanqlionu kapsuldaxili postkapilyar venulaların mikroskopik (A), onların divarının təşkilində iştirak edən strukturların ultrastruktur (B, C, Ç) şəkilləri. A yarımnazik kəsik, B, C, Ç elektronoqrammalar. İzahı mətndə verilmişdir. Boyaq: A metilen abısı / azur II + qələvi fuksin, B, C, Ç uranil asetat və təmiz qurğuşun sitrat. Miqyas A 20 mkm, B, C, Ç 500 nm.
Şək. 4.38. Onurğa beyni qanqlionunda kapsuldaxili damarların endotel örtüklərində formalaşan transendotel kanalı (A) və endotel hüceyrələri arasında (B, C, Ç) əlaqə formalarının elektron mikroskopik şəkilləri. TEM. İzahı mətndə verilmişdir. Boyaq: uranil asetat və təmiz qurğuşun sitrat. Böyütmə: A, C, Ç miqyas 200 nm, ) 42000 dəfə.
Sıx əlaqələr kapsul daxili postkapilyar venulalara məxsus endoteliositlər arasında da müşahidə olunur (şək. 4.38 ç-də oxlarla göstərilib). Ancaq qeyd etmək lazımdır ki, venulalarda endotel hüceyrələri arasında formalaşan sıx əlaqələrdəki bitişmə nöqtələri sayca xeyli az olur, bu isə onların keçiricilik qabiliyyətlərinin arteriolalara nisbətən artıq olduğunun ultrastruktur göstəricisi sayıla bilər.
MIKROQ@LIYA
Məlum olduğu kimi, sinir sisteminin makrofaqları hesab edilən mikroqliya hüceyrələri ölçülərinə görə sinir toxumasının ən kiçik hüceyrə qrupunu təşkil edir.
Qeyd etmək istərdik ki, beyin qabığında mikroqliya hüceyrələrinin müəyyən edilməsi əsas etibarilə Iba1 adlandırılan poliklonal əkscisimciklərin tətbiqi ilə aparılmışdır [199]. Belə ki, Iba1 yalnız mikroqliya hüceyrələrində ekspressiya olunan ionlaşmış kalsium-birləşdirici adapter molekullarını tanıyan əkscisimcikdir. Bu səbəbdən də tədqiqatımız zamanı beyin qabığı mikroqliyasını digər elementlərdən ayırd etmək üçün göstərilən işlərin nəticələrindən istifadə etmişik.
Nəzarət heyvan qrupundan əldə edilmiş histoloji kəsiklərdə müəyyən edirik ki, say etibarilə bu hüceyrələr beyin qabığının ən azsaylı hüceyrə kateqoriyasını təşkil edir. Elektronoqramlarda mikroqliositlərin cisimləri makrofaqlar üçün xarakterik olan oval (şək. 3.12a və 3.12b), uzunsov və ya üçbucaq formada nəzərə çarpır və yaxınlıqda yerləşən astrositin girintili-çıxıntılı və qeyri-müəyyən formaya malik (şək. 3.12b) hüceyrə cismindən fərqlənir. Mikroqliositlərin elektron sıxlığı OLD-in sıxlığından az olub sitoplazmasında OLD-lə müqayisədə daha az miqdarda ribosomlara və mikroborucuqlara təsadüf edilir.
Mikroqlial hüceyrələrin nüvəsində xromatinin kondensasiya dərəcəsi də astrositlər və oliqodendrositlərlə müqayisədə ara mövqe tutur (şək. 3.11 və 3.12). Belə ki, nüvə örtüyünün bilavasitə altında tünd təbəqə şəklində yerləşən heteroxromatinin qalınlığı astrositlərdə olduğundan qalın, oliqodendrositlərdəkindən isə nazikdir.
Nəzarət heyvanlardan alınmış preparatların təhlilini yekulaşdıraraq vurğulamaq istərdik ki, beyin qabığında müşahidə etdiyimiz mikroqliya hüceyrələrinin miqdarınım az, cisimlərinin kiçik, şaxələrinin isə nazik olması morfologiyası immunoloji cəhətdən sakit şaxələnmiş mikroqliya morfologiyasına tam uyğundur [117]. İstər yarımnazik, istərsə də ultranazik kəsiklərdə mikroqliya ilə təmasda yerləşmiş qabıq neyronlarının strukturunda heç bir patoloji dəyişiklik müşahidə edilmir (şək. 3.11 və şək.3.12).
Şək. 3.10. Beyin qabığı oliqodendrositlərinin ultrastrukturu. İzahı mətndə verilib. Miqyas a) 2mkm, b) 500nm.
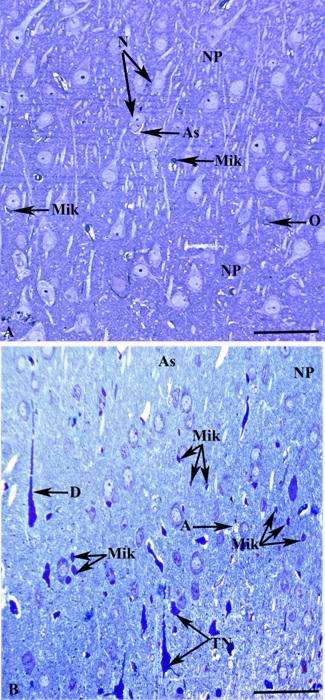
Şək. 3.11. Beyin qabığı parenximasının tərkibində yerləşən mikroqliya hüceyrələrinin ətraf elementlərlə qarşılıqlı histotopoqrafiyasının işıq mikroskopik şəkilləri. Boyaq toluidin abısı. İzahı mətndə verilib. Miqyas 20mkm.
Şək. 3.12. Beyin qabığında mikroqliya hüceyrələrinin ətraf elementlərlə qarşılıqlı histotopoqrafiyasının elektron mikroskopik şəkilləri. İzahı mətndə verilib. Miqyas: a) 10mkm, b) 2mkm.
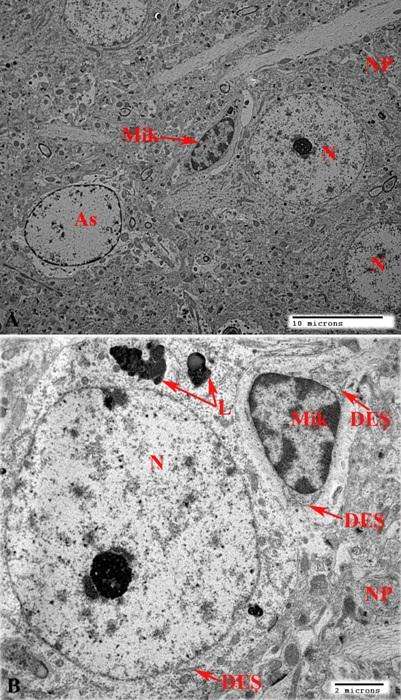
Nəzarət heyvanların beyin qabığı kəsikləri (şək. 3.22a) ilə kəskin endotoksemiya zamanı əldə edilmiş yarımnazik kəsiklərini müqayisə etdikdə ikincilərdə xüsusilə diqqəti cəlb edən əlamətlərdən biri də mikroqliya hüceyrələrinin miqdarının artmasıdır (şək. 3.22b). Bu hüceyrələrə beyin qabığının bütün qatlarında rast gəlmək mümkündür. Miqdarca daha çox nəzərə çarpan mikroqliositlər tək-tək yerləşməklə yanaşı həm də tünd destruktiv neyronların yaxınlığında toplantı şəklində nəzərə çarpırlar (şək. 3.22b). Əgər nəzarət kəsiklərdə (şək 3.11b, 3.22a) mikroqliya ilə bilavasitə əlaqədə olan neyronlarda morfoloji cəhətdən heç bir patoloji dəyişiklik müşahidə edilmirsə, şəkil 22b- dən göründüyü kimi endotoksemiya zamanı istər mikroqliya ilə təmasda olan, eləcə də ətrafda yerləşən sinir hüceyrələrində degenerasiya əlamətləri aşkarlanır. Digər bir tərəfdən, endotoksemiya zamanı degenerativ neyronlar əksər hallarda mikroqliositlərin nisbətən çox olduğu nahiyyələrində rast gəlinir (şək. 3.22b). Belə ki, şəkil 3.22b – də təsvir edilmiş yarımnazik kəsikdən güründüyü kimi bir sinir hüceyrəsi ilə təmasda üç mikroqliya hüceyrəsi müəyyən olunur. Həmin neyron normal strukturlu neyronlara nisbətən tünd, artıq degenerasiyaya uğramış tünd neyronlarla müqayisədə isə açıq rəngə boyanmışdır. Bu göstərici qeyd olunan neyronda degenerativ proseslərin başlandığını və mikroqliyanın bu prosesdə bilavasitə iştirak etdiyini nümayiş etdirir.
Mikroqliositlərin miqdarının damarətrafı zonalarda da artması, eləcə də endotoksemiya zamanı bu hüceyrələrin ultrastruktur dəyişiklikləri beyin toxumasında iltihab reaksiyalarının inkişaf etdiyini birmənalı şəkildə təsdiq edir (şək. 3.22b, 3.23).
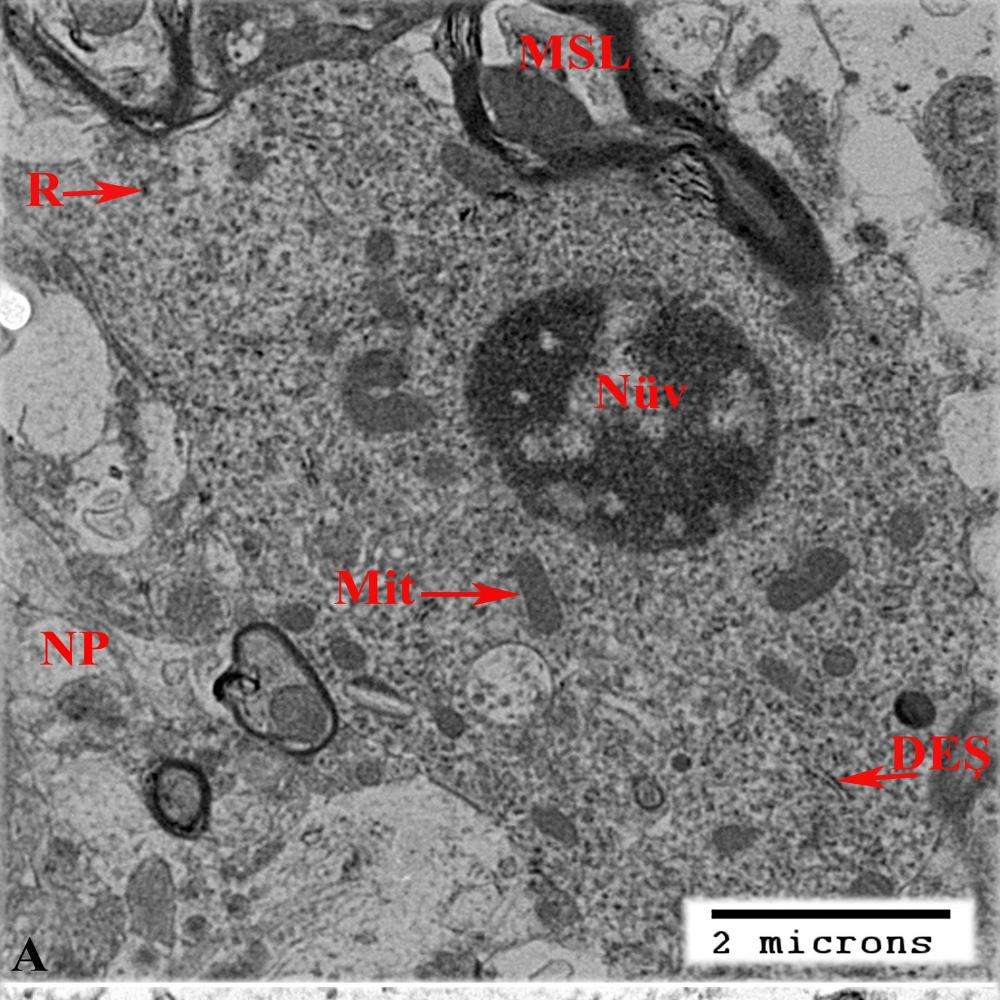
Qeyd etmək lazımdır ki, kəskin endotoksemiya zamanı hazırlanmış preparatlarda mikroqliyanın morfoloji cəhətdən müxtəlifliyi bu hüceyrələrdə venaya yeridilmiş lipopolisaxaridə qarşı yönəlmiş cavab reaksiyanın müxtəlif mərhələlərini izləməyə imkan yaradır. Başqa sözlə, müşahidə olunan morfoloji tiplər əslində bir-birini ardıcıl olaraq əvəz edir. Şəkil 3.23a – da nümayiş etdilmiş mikrofotoda degenerasiyaya uğrayan neyronla bilavasitə təmasda bütün ultrastruktur göstəricilərinə görə mikroqliyaya tam uyğun olan hüceyrədə sərbəst ribosomların miqdarının nəzərə çarpacaq dərəcədə artmasını müşahidə edirik. Göstərilən əlamətlər endotoksemiyanın ilk mərhələlərində mikroqliyada sintetik aktivliyin güclənməsini nümayiş etdirir. Şəkil 3.23b-də isə prekapilyar arterioladan kapilyarın başlanğıc götürdüyü nahiyyənin bilavasitə yaxınlığında yerləşən mikroqliya hüceyrəsi nümayiş etdirilmişdir. Qeyd olunan damarlar ilə mikroqliya hüceyrəsi arasında astrosit ayaqcığından başqa digər törəmənin müşahidə edilməməsi, digər tərəfdən isə təsvir olunan kapilyarın mənfəzinin qanın formalı elementi olan monositlə tutulması şəkil 23b – də xüsusilə diqqəti cəlb edən məqamlardandır.
Şək. 3.22. Kəskin endotoksemiya zamanı beyin qabığı mikroqliya hüceyrələrində baş verən dəyişikliklərin işıq mikroskopik şəkilləri. İzahı mətndə verilib. Boyaq metilen abısı. Miqyas: 40mkm.
Şək. 3.23. Kəskin endotoksemiya zamanı beyin qabığı mikroqliya hüceyrələrində və ətraf törəmələrdə baş verən dəyişikliklərin elektron mikroskopik şəkilləri. İzahı mətndə verilib. Miqyas: a) və b) 2mkm; c) başlanğıc böy. – x5000.
Mikroqliositin sitoplazması şəkil 23a – da nümayiş etdirilmiş mikroqliositlə müqayisədə nəzərə çarpacaq dərəcədə tündləşmişdir. Sitoplazmada istər sərbəst, istərsə də dənəli endoplazmatik şəbəkə ilə əlaqədali ribosomların miqdarı artmışdır ki, bu da perivaskulyar mikroqliyada da zülal sintezinin aktivləşməsi kimi qiymətləndirilməlidir.
Şəkil 3.23c –də isə degenerativ sinir hüceyrəsi ilə təmasda iki bir-birinə söykənmiş mikroqliya hüceyrəsinin sitoplazmasının tündləşməsi qeyd olunur (şək. 3.23c). Hüceyrələrin nüvələrinin ölçüləri böyümüş, nəticədə nüvə-sitoplazma nisbəti yüksəlmişdir (şək. 3.23c). İçərisində ayrı-ayrı ribosomlar müşahidə edilən sitoplazma nazik halqa şəklində nüvəni əhatə etmişdir.
Perivaskulyar mikroqliyada baş verən sonrakı dəyişikliklər şəkil 3.24 a -da nümayiş etdirilmişdir. Burada kəskin iltihab nəticəsində perivaskulyar mikroqliyada xromatinin kondensasiya dərəcəsinin artması qeyd olunur. Damar mənfəzində miyelinə bənzər cisimcik, endotel plazmalemmasının isə tamlığının pozulması müəyyən edilir. Bütün bunlar, eləcə də ətraf neyropildə və astrosit ayaqcıqlarında ödem mayesinin müşahidə edilməsi damar keçiriciliyinin artdığını nümayiş etdirir. Neyropil elementləri arasında şəkil 3.24a –da göy oxlarla işarə edilmiş və vazogen mənşəli ödemə müvafiq gələn elektron mikroskopik cəhətdən açıq hissələr qeyd olunur ki, oxşar görüntülər nəzarət preparatlarda rast gəlinmir. Ən maraqlısı isə ondan ibarətdir ki, ödem daha çox aktivləşmiş mikroqliyanın yerləşdiyi tərəfdə nəzərə çarpır (şək. 3.24a-da sol tərəfdə).
Bu hal, eləcə də mikroqliya hüceyrəsinin qan damarı ilə şişkinləşmiş astrosit ayaqcığı arasında yerləşməsi mikroqliayanın beyin ödeminin yaranmasındakı bilavasitə rolu barədə məlumat verir. Mikroqliya sitoplazmasında pinositoz qovuqcuqların, birincili və ikincili lizosomların və multivezikulyar cisimciklərin miqdarı artmışdır. Göstərilən əlamətlər mikroqliyanın pinosotoz aktivliyinin artdığını vurğulayır (şək. 3.24a). Şəkil 3.24b - də nümayiş olunmuş elektronnoqramlarda isə əvvəlki şəkillərlə müqayisədə sitoplazması xeyli tündləşmiş mikroqliositlər nəzərə çarpır.
Şək. 3.24. Kəskin endotoksemiya zamanı perivaskulyar (a) və neyropil daxilində yerləşən mikroqliya hüceyrələrinin ultrastrukturu. İzahı mətndə verilib. Miqyas: 2mkm.
Sitoplazmanın lizosomlar, eləcə də müxtəlif formaya malik faqolizosomlarla dolu olması isə kəskin endotoksemiya zamanı mikroqliyada faqositar aktivliyin də yüksəldiyini birmənalı şəkildə sübut edir (şək. 3.24b). Ətraf toxumada yerləşən sinir hüceyrələrində, eləcə də miyelinli sinir liflərində baş vermiş destruktiv dəyişikliklər də nəzər diqqəti cəlb edir (şək. 3.24b).
Tədqiqat apardığımız heyvanların beyin qabıqlarından hazırlanmış kəsiklərdən aydın olur ki, mikroqliositlərin degenerativ neyronlarla təmasda yerləşməsi son nəticədə bu hüceyrələrin özlərində də müəyyən destruktiv əlamətlərin meydana çıxmasına səbəb olur (şək. 3.25). Şəkil 3.25a – da degenerasiya uğramış neyronla əlaqədə olan, şəkil 3.25b – də isə bir qədər məsafədə yerləşmiş mikroqliyanın destruksiyasını nümayiş etdirən elektronnoqramlar verilmişdir. Belə ki, qeyd olunan hüceyrələrdə sitoplazmatik elementlərə aid yalnız şişkinləşmiş destruktiv mitoxondriləri ayırd etmək mümkündür. Hüceyrə möhtəviyyatının destruksiyaya uğraması nəticəsində sitoplazma homogen görünüş almışdır.
Beləliklə, tədqiqatın nəticələrindən göründüyü kimi, kəskin endotoksemiya zamanı beyin qabığının sinir və damar elementlərində müşahidə edilən müxtəlif dərəcəli mexaniki və morfofunksional dəyişikliklərdə aktiv iştirak edən mikroqliya hüceyrələrinin özləri də, son nəticədə, əsaslı patoloji dəyişikliklərə uğrayır.